
Boundary Maps
Here’s a brief recap of the previous article: we learnt that in refining a cell decomposition of an object M, we can, at each step, pick an i-dimensional cell and divide it in two. In this way, we introduce an additional i-dimensional cell and an (i-1)-dimensional cell.
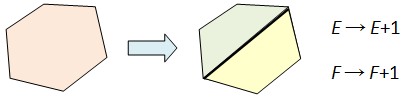
Hence even after successive steps, the Euler characteristic 
In fact, it is possible to obtain even finer distinguishing characteristics for the topology of M, known as Betti numbers. For starters, consider the boundary function ∂ which takes a cell to its boundary ∂C, while taking into account its orientation. If C is i-dimensional, then ∂C is a union of (i-1)-dimensional cells. For example, in the following diagram, ∂C can be expressed as a sum of 5 edges:

The critical observation is that when we partition a cell into two, written as 
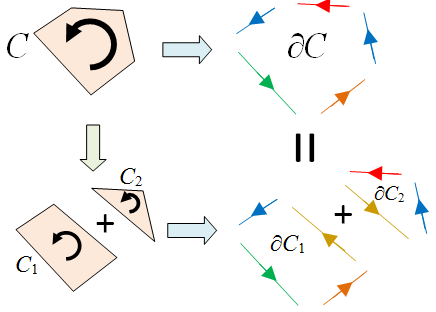
since the two edges which are equal but in opposite directions cancel each other out. The next property we note is that ∂(∂C) = 0, as illustrated by the following diagram.
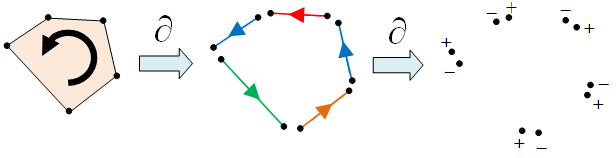
Betti Numbers
This suggests the following: let Si denote the set of i-dimensional cells in the decomposition. An i–chain is defined as a formal sum 
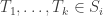


- addition:
;
- scalar multiplication:
.
The boundary map then gives a linear map:
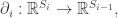
such that the composition 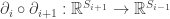
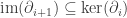

Definition. The i-th Betti number, denoted bi, is the dimension of the quotient
.
We’ll briefly justify why this definition is independent of our choice of cell decomposition. It suffices to reduce to the case where we partition an i-dimensional cell

The boundary maps are then modified by adding the following relations:

from which we can show that 

Sample Computation: Torus
Consider the torus, which is obtained by gluing the opposite sides of a square together in the same direction:
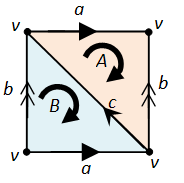
The above cellular decomposition then comprises of the following cells:
- 2-cells : {A, B}, oriented in clockwise;
- 1-cells : {a, b, c}, with orientation as above;
- 0-cells : {v}, since all four vertices of the square are glued together.
Then we have the following relations:
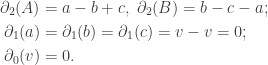
Now we can easily compute the Betti numbers of the torus.
- First,
is spanned by a single element A+B so it has dimension 1. Thus
- Next,
is spanned by a+c–b and has dimension 1. And
is the full space spanned by {a, b, c}, so
- Finally,
and
is spanned by v and has dimension 1, so
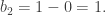
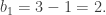
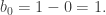
Exercise
Verify the Betti numbers of the following surfaces:


Homology Groups
In fact, by considering the abelian groups 

For example, consider the projective plane and the square which have the same Betti numbers.
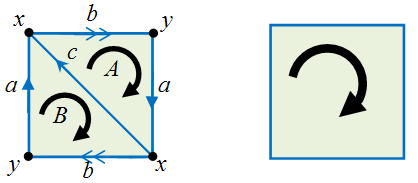
If we let M be the projective plane on the left, we get:
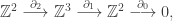

We still have 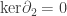
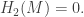





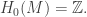
On the other hand, we leave it to the reader to check that if N is the square, then 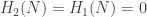

Exercise
Compute the homology groups for each of the nine shapes in the previous exercise.
Simplicial Homology
At higher dimensions, it will be a hassle to illustrate the various cells and their boundaries. Thankfully, in the case of simplicial complexes, this can be calculated combinatorically without any need for diagrams. Here, each m-dimensional cell is written as an (m+1)-tuple of points: ![\Delta = [ v_0, v_1, \ldots, v_m ]](https://s0.wp.com/latex.php?latex=%5CDelta+%3D+%5B+v_0%2C+v_1%2C+%5Cldots%2C+v_m+%5D&bg=ffffff&fg=333333&s=0&c=20201002)
![\partial_i([v_0, v_1, \ldots, v_m]) := \sum_{i=0}^m (-1)^i [v_0, \ldots, \hat{v_i}, \ldots, v_m],](https://s0.wp.com/latex.php?latex=%5Cpartial_i%28%5Bv_0%2C+v_1%2C+%5Cldots%2C+v_m%5D%29+%3A%3D+%5Csum_%7Bi%3D0%7D%5Em+%28-1%29%5Ei+%5Bv_0%2C+%5Cldots%2C+%5Chat%7Bv_i%7D%2C+%5Cldots%2C+v_m%5D%2C&bg=ffffff&fg=333333&s=0&c=20201002)
where the i-th term has all vertices except 
![\partial_2([a, b, c]) = [b,c] - [a,c] + [a,b].](https://s0.wp.com/latex.php?latex=%5Cpartial_2%28%5Ba%2C+b%2C+c%5D%29+%3D+%5Bb%2Cc%5D+-+%5Ba%2Cc%5D+%2B+%5Ba%2Cb%5D.&bg=ffffff&fg=333333&s=0&c=20201002)

Example
Suppose we have the following simplicial complex which is the union of a tetrahedron’s surface and a triangle.
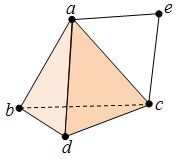
This comprises of four 2-simplices, eight 1-simplices and five 0-simplices. The boundary maps are given as above, e.g. ![\partial_2([a, c, d]) = [c,d] - [a,d] + [a,c],](https://s0.wp.com/latex.php?latex=%5Cpartial_2%28%5Ba%2C+c%2C+d%5D%29+%3D+%5Bc%2Cd%5D+-+%5Ba%2Cd%5D+%2B+%5Ba%2Cc%5D%2C&bg=ffffff&fg=333333&s=0&c=20201002)
![\partial_1([a,e]) = [e] - [a]](https://s0.wp.com/latex.php?latex=%5Cpartial_1%28%5Ba%2Ce%5D%29+%3D+%5Be%5D+-+%5Ba%5D&bg=ffffff&fg=333333&s=0&c=20201002)
![\partial_0([d]) = 0.](https://s0.wp.com/latex.php?latex=%5Cpartial_0%28%5Bd%5D%29+%3D+0.&bg=ffffff&fg=333333&s=0&c=20201002)

In general, expressing a topological object as a simplicial complex can be a rather tedious affair, for the vertices of each simplex must be distinct, and two distinct simplices cannot be represented by the same tuple of vertices. E.g. in the case of a torus, one possibility is the following:
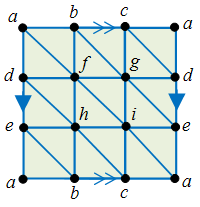
which has 18 faces, 27 edges and 9 vertices. Hence, simplicial homology can be quite a hassle to compute by hand, but on the plus side one can easily write a program to compute it. Also, note that simplicial homology is a special case of cellular homology (one where all cells are simplices).
Exercise. What’s wrong with the following simplicial complex for the torus?
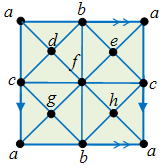 Answer (highlight to read) : the 1-simplices [a, c] and [a, b] are duplicated. E.g. [a, b] appears twice for the two edges at the bottom of the square.
Answer (highlight to read) : the 1-simplices [a, c] and [a, b] are duplicated. E.g. [a, b] appears twice for the two edges at the bottom of the square.
Betti Numbers and Euler Characteristic
One can obtain the Euler characteristic of M from its Betti numbers:
Theorem. For any object M, we have:
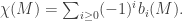
Proof
We know from linear algebra that for any linear map T : V → W, we have dim(ker(T)) + dim(im(T)) = dim(V). Hence for each i, we have
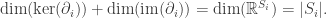
And since 
![\begin{aligned}\sum_i (-1)^i b_i(M) &= \sum_i (-1)^i\dim(\text{ker}(\partial_i)) - (-1)^i\sum_i\dim(\text{im}(\partial_{i+1}))\\ &=\sum_i (-1)^i \dim(\text{ker} (\partial_i)) + \sum_i (-1)^{i+1}\dim( \text{im}(\partial_{i+1})) \\ &=\sum_i (-1)^i [\dim(\text{ker} (\partial_i)) + \dim(\text{im}(\partial_i))] \\ &= \sum_i (-1)^i |S_i| =\chi(M)\end{aligned}](https://s0.wp.com/latex.php?latex=%5Cbegin%7Baligned%7D%5Csum_i+%28-1%29%5Ei+b_i%28M%29+%26%3D+%5Csum_i+%28-1%29%5Ei%5Cdim%28%5Ctext%7Bker%7D%28%5Cpartial_i%29%29+-+%28-1%29%5Ei%5Csum_i%5Cdim%28%5Ctext%7Bim%7D%28%5Cpartial_%7Bi%2B1%7D%29%29%5C%5C+%26%3D%5Csum_i+%28-1%29%5Ei+%5Cdim%28%5Ctext%7Bker%7D+%28%5Cpartial_i%29%29+%2B+%5Csum_i+%28-1%29%5E%7Bi%2B1%7D%5Cdim%28+%5Ctext%7Bim%7D%28%5Cpartial_%7Bi%2B1%7D%29%29+%5C%5C+%26%3D%5Csum_i+%28-1%29%5Ei+%5B%5Cdim%28%5Ctext%7Bker%7D+%28%5Cpartial_i%29%29+%2B+%5Cdim%28%5Ctext%7Bim%7D%28%5Cpartial_i%29%29%5D+%5C%5C+%26%3D+%5Csum_i+%28-1%29%5Ei+%7CS_i%7C+%3D%5Cchi%28M%29%5Cend%7Baligned%7D&bg=ffffff&fg=333333&s=0&c=20201002)
as desired. ♦
Mayer-Vietoris Sequence
Suppose 


[ Note: an exact sequence of abelian groups is a sequence of maps as above such that for any two consecutive maps 

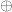
Let’s explain the various maps. First, pick a cellular decomposition of M and of N; then find a decomposition of X which is a common refinement of both. The inclusion 





[ Note: even though 
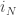
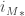





as the map
,
as the map
![[D] \mapsto {i_M}_*([D]) + {i_N}_*([D]).](https://s0.wp.com/latex.php?latex=%5BD%5D+%5Cmapsto+%7Bi_M%7D_%2A%28%5BD%5D%29+%2B+%7Bi_N%7D_%2A%28%5BD%5D%29.&bg=ffffff&fg=333333&s=0&c=20201002)
The map left of interest is the boundary map:
For that, suppose we have an m-chain D of 
[ For the conscientious reader, the map is well-defined because
- ∂(∂D’) = 0 so we can take the class of ∂D’ in M ∩ N;
- if we replace D by D+∂E, we can write E as a sum of (m+1)-chains E’+E”, where E’, E” are chains of M, N respectively, so D’ is replaced by D’+∂E’ which gives ∂D’ = ∂(D’+∂E’) = ∂D’;
- if we pick a different D = D1‘ + D2“, then D1‘–D’ = D”–D2“ is a chain in M ∩ N, and thus ∂D1‘ = ∂(D1‘–D’) + ∂D’ has the same image as ∂D’ modulo im(∂) in M ∩ N. ]
Example
Let consider the 3-sphere 
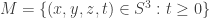
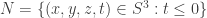
with intersection 

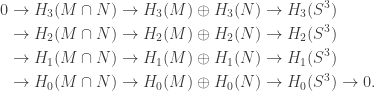
Since M and N are homeomorphic to the solid ball 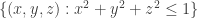

and 


Hi there! Just wanted to ask about the definition of chains. Here you use formal linear combinations of simplices as a chain, but in Calculus on Manifolds by Spivak, he uses formal linear combinations of (what he calls) singular n-cubes, which are maps from [0,1]^n –> R^s. I know that you should be able to integrate on both kinds of chains simply, but does one offer any advantage over the other? Why is there a difference?
The difference lies in singular or simplicial homology. Spivak is talking about singular homology while I’m talking about simplicial.
Roughly, in singular homology you can prove things but can’t really compute; in simplicial homology you can compute but can’t prove much.The tough part is to show that the two are equal on a huge class of topological spaces (e.g. CW-complexes).